

What Is Base Pair Rule

A base of operations pair (bp) is a primal unit of double-stranded nucleic acids consisting of two nucleobases bound to each other past hydrogen bonds. They form the building blocks of the DNA double helix and contribute to the folded structure of both DNA and RNA. Dictated past specific hydrogen bonding patterns, "Watson–Crick" (or "Watson–Crick–Franklin") base of operations pairs (guanine–cytosine and adenine–thymine)[i] allow the DNA helix to maintain a regular helical structure that is subtly dependent on its nucleotide sequence.[two] The complementary nature of this based-paired structure provides a redundant re-create of the genetic information encoded within each strand of Dna. The regular structure and data redundancy provided by the DNA double helix make DNA well suited to the storage of genetic information, while base-pairing betwixt DNA and incoming nucleotides provides the machinery through which DNA polymerase replicates DNA and RNA polymerase transcribes DNA into RNA. Many Dna-binding proteins can recognize specific base-pairing patterns that identify particular regulatory regions of genes.
Intramolecular base pairs can occur inside unmarried-stranded nucleic acids. This is particularly important in RNA molecules (east.grand., transfer RNA), where Watson–Crick base of operations pairs (guanine–cytosine and adenine–uracil) permit the formation of curt double-stranded helices, and a wide diverseness of not–Watson–Crick interactions (e.g., Yard–U or A–A) allow RNAs to fold into a vast range of specific 3-dimensional structures. In improver, base of operations-pairing between transfer RNA (tRNA) and messenger RNA (mRNA) forms the footing for the molecular recognition events that upshot in the nucleotide sequence of mRNA becoming translated into the amino acrid sequence of proteins via the genetic code.
The size of an individual gene or an organism's entire genome is often measured in base pairs because Deoxyribonucleic acid is usually double-stranded. Hence, the number of total base pairs is equal to the number of nucleotides in one of the strands (with the exception of non-coding single-stranded regions of telomeres). The haploid man genome (23 chromosomes) is estimated to be near 3.two billion bases long and to contain 20,000–25,000 distinct protein-coding genes.[3] [iv] [five] A kilobase (kb) is a unit of measurement in molecular biology equal to chiliad base pairs of DNA or RNA.[6] The total number of Dna base pairs on World is estimated at 5.0×x 37 with a weight of 50 billion tonnes.[7] In comparison, the total mass of the biosphere has been estimated to be as much equally 4 TtC (trillion tons of carbon).[8]
Hydrogen bonding and stability [edit]


Top, a Grand.C base pair with three hydrogen bonds. Bottom, an A.T base pair with two hydrogen bonds. Non-covalent hydrogen bonds betwixt the bases are shown equally dashed lines. The wiggly lines stand up for the connexion to the pentose sugar and bespeak in the direction of the minor groove.
Hydrogen bonding is the chemical interaction that underlies the base-pairing rules described above. Advisable geometrical correspondence of hydrogen bond donors and acceptors allows only the "correct" pairs to form stably. Dna with high GC-content is more than stable than DNA with low GC-content. Only, reverse to popular belief, the hydrogen bonds do not stabilize the DNA significantly; stabilization is mainly due to stacking interactions.[9]
The bigger nucleobases, adenine and guanine, are members of a class of double-ringed chemic structures called purines; the smaller nucleobases, cytosine and thymine (and uracil), are members of a class of single-ringed chemic structures called pyrimidines. Purines are complementary only with pyrimidines: pyrimidine-pyrimidine pairings are energetically unfavorable considering the molecules are too far apart for hydrogen bonding to be established; purine-purine pairings are energetically unfavorable considering the molecules are too close, leading to overlap repulsion. Purine-pyrimidine base-pairing of AT or GC or UA (in RNA) results in proper duplex structure. The merely other purine-pyrimidine pairings would be Air-conditioning and GT and UG (in RNA); these pairings are mismatches because the patterns of hydrogen donors and acceptors do not correspond. The GU pairing, with two hydrogen bonds, does occur fairly often in RNA (come across wobble base pair).
Paired Dna and RNA molecules are comparatively stable at room temperature, only the two nucleotide strands will separate above a melting signal that is adamant past the length of the molecules, the extent of mispairing (if whatever), and the GC content. Higher GC content results in college melting temperatures; it is, therefore, unsurprising that the genomes of extremophile organisms such as Thermus thermophilus are specially GC-rich. On the converse, regions of a genome that demand to divide oftentimes — for instance, the promoter regions for often-transcribed genes — are comparatively GC-poor (for example, run into TATA box). GC content and melting temperature must also be taken into account when designing primers for PCR reactions.[ commendation needed ]
Examples [edit]
The post-obit DNA sequences illustrate pair double-stranded patterns. By convention, the top strand is written from the five′-cease to the 3′-end; thus, the bottom strand is written 3′ to 5′.
- A base of operations-paired Deoxyribonucleic acid sequence:
-
ATCGATTGAGCTCTAGCG -
TAGCTAACTCGAGATCGC
-
- The corresponding RNA sequence, in which uracil is substituted for thymine in the RNA strand:
-
AUCGAUUGAGCUCUAGCG -
UAGCUAACUCGAGAUCGC
-
Base of operations analogs and intercalators [edit]
Chemic analogs of nucleotides can have the place of proper nucleotides and institute non-canonical base-pairing, leading to errors (mostly betoken mutations) in DNA replication and Dna transcription. This is due to their isosteric chemistry. One common mutagenic base of operations analog is 5-bromouracil, which resembles thymine just can base of operations-pair to guanine in its enol form.[ citation needed ]
Other chemicals, known as Deoxyribonucleic acid intercalators, fit into the gap between adjacent bases on a single strand and induce frameshift mutations past "masquerading" as a base, causing the Dna replication mechanism to skip or insert additional nucleotides at the intercalated site. Most intercalators are large polyaromatic compounds and are known or suspected carcinogens. Examples include ethidium bromide and acridine.[ commendation needed ]
Mismatch repair [edit]
Mismatched base of operations pairs can be generated by errors of DNA replication and as intermediates during homologous recombination. The process of mismatch repair usually must recognize and correctly repair a small number of base mispairs within a long sequence of normal DNA base pairs. To repair mismatches formed during Deoxyribonucleic acid replication, several distinctive repair processes have evolved to distinguish betwixt the template strand and the newly formed strand and then that only the newly inserted incorrect nucleotide is removed (in order to avert generating a mutation).[x] The proteins employed in mismatch repair during DNA replication, and the clinical significance of defects in this process are described in the article Dna mismatch repair. The process of mispair correction during recombination is described in the article cistron conversion.
Unnatural base of operations pair (UBP) [edit]
An unnatural base pair (UBP) is a designed subunit (or nucleobase) of Dna which is created in a laboratory and does non occur in nature. DNA sequences have been described which utilise newly created nucleobases to form a third base of operations pair, in addition to the two base pairs found in nature, A-T (adenine – thymine) and G-C (guanine – cytosine). A few inquiry groups accept been searching for a third base of operations pair for Dna, including teams led by Steven A. Benner, Philippe Marliere, Floyd Due east. Romesberg and Ichiro Hirao.[11] Some new base of operations pairs based on culling hydrogen bonding, hydrophobic interactions and metal coordination have been reported.[12] [13] [14] [15]
In 1989 Steven Benner (then working at the Swiss Federal Establish of Technology in Zurich) and his team led with modified forms of cytosine and guanine into DNA molecules in vitro.[16] The nucleotides, which encoded RNA and proteins, were successfully replicated in vitro. Since then, Benner's team has been trying to engineer cells that can make foreign bases from scratch, obviating the need for a feedstock.[17]
In 2002, Ichiro Hirao's group in Nihon developed an unnatural base of operations pair between 2-amino-8-(2-thienyl)purine (s) and pyridine-2-one (y) that functions in transcription and translation, for the site-specific incorporation of non-standard amino acids into proteins.[18] In 2006, they created 7-(ii-thienyl)imidazo[iv,5-b]pyridine (Ds) and pyrrole-two-carbaldehyde (Pa) as a third base pair for replication and transcription.[19] Afterward, Ds and iv-[3-(six-aminohexanamido)-one-propynyl]-2-nitropyrrole (Px) was discovered as a high fidelity pair in PCR amplification.[20] [21] In 2013, they applied the Ds-Px pair to Deoxyribonucleic acid aptamer generation by in vitro selection (SELEX) and demonstrated the genetic alphabet expansion significantly augment DNA aptamer affinities to target proteins.[22]
In 2012, a grouping of American scientists led by Floyd Romesberg, a chemical biologist at the Scripps Research Institute in San Diego, California, published that his team designed an unnatural base pair (UBP).[xiv] The 2 new artificial nucleotides or Unnatural Base of operations Pair (UBP) were named d5SICS and dNaM. More technically, these artificial nucleotides bearing hydrophobic nucleobases, characteristic two fused aromatic rings that form a (d5SICS–dNaM) complex or base of operations pair in DNA.[17] [23] His team designed a variety of in vitro or "test tube" templates containing the unnatural base pair and they confirmed that it was efficiently replicated with high fidelity in near all sequence contexts using the modern standard in vitro techniques, namely PCR amplification of Deoxyribonucleic acid and PCR-based applications.[fourteen] Their results show that for PCR and PCR-based applications, the d5SICS–dNaM unnatural base pair is functionally equivalent to a natural base pair, and when combined with the other ii natural base of operations pairs used by all organisms, A–T and One thousand–C, they provide a fully functional and expanded six-alphabetic character "genetic alphabet".[23]
In 2014 the same team from the Scripps Enquiry Institute reported that they synthesized a stretch of circular DNA known as a plasmid containing natural T-A and C-G base pairs along with the all-time-performing UBP Romesberg's laboratory had designed and inserted it into cells of the common bacterium E. coli that successfully replicated the unnatural base pairs through multiple generations.[11] The transfection did not hamper the growth of the East. coli cells and showed no sign of losing its unnatural base pairs to its natural Dna repair mechanisms. This is the offset known example of a living organism passing along an expanded genetic code to subsequent generations.[23] [24] Romesberg said he and his colleagues created 300 variants to refine the blueprint of nucleotides that would exist stable enough and would be replicated as easily equally the natural ones when the cells divide. This was in part achieved by the addition of a supportive algal gene that expresses a nucleotide triphosphate transporter which efficiently imports the triphosphates of both d5SICSTP and dNaMTP into E. coli bacteria.[23] Then, the natural bacterial replication pathways utilize them to accurately replicate a plasmid containing d5SICS–dNaM. Other researchers were surprised that the bacteria replicated these human-fabricated Dna subunits.[25]
The successful incorporation of a third base pair is a significant breakthrough toward the goal of greatly expanding the number of amino acids which tin be encoded past Deoxyribonucleic acid, from the existing 20 amino acids to a theoretically possible 172, thereby expanding the potential for living organisms to produce novel proteins.[11] The artificial strings of Dna do not encode for anything however, only scientists speculate they could exist designed to manufacture new proteins which could have industrial or pharmaceutical uses.[26] Experts said the synthetic DNA incorporating the unnatural base of operations pair raises the possibility of life forms based on a different DNA code.[25] [26]
Non-approved base pairing [edit]
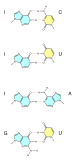
Wobble base pairs
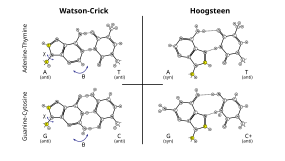
Comparison of Hoogsteen to Watson–Crick base pairs.[27]
In add-on to the approved pairing, some weather can also favour base-pairing with alternative base orientation, and number and geometry of hydrogen bonds. These pairings are accompanied by alterations to the local backbone shape.[ citation needed ]
The nigh common of these is the wobble base pairing that occurs between tRNAs and mRNAs at the third base position of many codons during transcription[28] and during the charging of tRNAs past some tRNA synthetases.[29] They have also been observed in the secondary structures of some RNA sequences.[30]
Additionally, Hoogsteen base pairing (typically written equally A•U/T and G•C) tin exist in some Dna sequences (e.g. CA and TA dinucleotides) in dynamic equilibrium with standard Watson–Crick pairing.[27] They accept too been observed in some poly peptide–DNA complexes.[31]
In addition to these culling base pairings, a wide range of base-base of operations hydrogen bonding is observed in RNA secondary and 3rd structure.[32] These bonds are often necessary for the precise, complex shape of an RNA, as well as its binding to interaction partners.[32]
Length measurements [edit]
The following abbreviations are normally used to describe the length of a D/RNA molecule:
- bp = base pair—1 bp corresponds to approximately 3.4 Å (340 pm)[33] of length along the strand, and to roughly 618 or 643 daltons for Dna and RNA respectively.
- kb (= kbp) = kilo–base of operations-pair = 1,000 bp
- Mb (= Mbp) = mega–base-pair = 1,000,000 bp
- Gb = giga–base-pair = ane,000,000,000 bp.
For single-stranded DNA/RNA, units of nucleotides are used—abbreviated nt (or knt, Mnt, Gnt)—every bit they are not paired. To distinguish between units of computer storage and bases, kbp, Mbp, Gbp, etc. may be used for base of operations pairs.
The centimorgan is as well frequently used to imply distance forth a chromosome, just the number of base pairs it corresponds to varies widely. In the human genome, the centimorgan is almost 1 meg base pairs.[34] [35]
See also [edit]
- List of Y-DNA unmarried-nucleotide polymorphisms
- Not-canonical base pairing
- Chargaff's rules
References [edit]
- ^ Spencer, Chiliad. (x January 1959). "The stereochemistry of deoxyribonucleic acid. II. Hydrogen-bonded pairs of bases". Acta Crystallographica. 12 (1): 66–71. doi:10.1107/S0365110X59000160. ISSN 0365-110X.
- ^ Zhurkin VB, Tolstorukov MY, Xu F, Colasanti AV, Olson WK (2005). "Sequence-Dependent Variability of B-DNA". Deoxyribonucleic acid Conformation and Transcription. pp. xviii–34. doi:10.1007/0-387-29148-2_2. ISBN978-0-387-25579-8.
- ^ Moran LA (2011-03-24). "The total size of the human genome is very likely to be ~3,200 Mb". Sandwalk.blogspot.com. Retrieved 2012-07-16 .
- ^ "The finished length of the human being genome is ii.86 Gb". Strategicgenomics.com. 2006-06-12. Retrieved 2012-07-16 .
- ^ International Human Genome Sequencing Consortium (Oct 2004). "Finishing the euchromatic sequence of the man genome". Nature. 431 (7011): 931–45. Bibcode:2004Natur.431..931H. doi:10.1038/nature03001. PMID 15496913.
- ^ Cockburn AF, Newkirk MJ, Firtel RA (December 1976). "Arrangement of the ribosomal RNA genes of Dictyostelium discoideum: mapping of the nontranscribed spacer regions". Prison cell. 9 (iv Pt one): 605–thirteen. doi:10.1016/0092-8674(76)90043-Ten. PMID 1034500. S2CID 31624366.
- ^ Nuwer R (18 July 2015). "Counting All the DNA on Earth". The New York Times. New York. ISSN 0362-4331. Archived from the original on 2022-01-01. Retrieved 2015-07-18 .
- ^ "The Biosphere: Diverseness of Life". Aspen Global Change Institute. Basalt, CO. Retrieved 2015-07-19 .
- ^ Yakovchuk P, Protozanova E, Frank-Kamenetskii Medico (2006-01-30). "Base of operations-stacking and base of operations-pairing contributions into thermal stability of the DNA double helix". Nucleic Acids Research. 34 (2): 564–74. doi:x.1093/nar/gkj454. PMC1360284. PMID 16449200.
- ^ Putnam, Christopher D. (September 2021). "Strand discrimination in Dna mismatch repair". DNA Repair. 105: 103161. doi:10.1016/j.dnarep.2021.103161. PMC8785607. PMID 34171627.
- ^ a b c Fikes BJ (May 8, 2014). "Life engineered with expanded genetic lawmaking". San Diego Union Tribune. Archived from the original on 9 May 2014. Retrieved 8 May 2014.
- ^ Yang Z, Chen F, Alvarado JB, Benner SA (September 2011). "Amplification, mutation, and sequencing of a vi-alphabetic character constructed genetic organisation". Journal of the American Chemic Society. 133 (38): 15105–12. doi:x.1021/ja204910n. PMC3427765. PMID 21842904.
- ^ Yamashige R, Kimoto M, Takezawa Y, Sato A, Mitsui T, Yokoyama S, Hirao I (March 2012). "Highly specific unnatural base pair systems equally a third base of operations pair for PCR amplification". Nucleic Acids Inquiry. twoscore (half dozen): 2793–806. doi:10.1093/nar/gkr1068. PMC3315302. PMID 22121213.
- ^ a b c Malyshev DA, Dhami K, Quach HT, Lavergne T, Ordoukhanian P, Torkamani A, Romesberg FE (July 2012). "Efficient and sequence-independent replication of DNA containing a third base pair establishes a functional six-letter genetic alphabet". Proceedings of the National University of Sciences of the Usa of America. 109 (30): 12005–10. Bibcode:2012PNAS..10912005M. doi:10.1073/pnas.1205176109. PMC3409741. PMID 22773812.
- ^ Takezawa, Yusuke; Müller, Jens; Shionoya, Mitsuhiko (2017-05-05). "Artificial DNA Base Pairing Mediated by Diverse Metal Ions". Chemistry Messages. 46 (5): 622–633. doi:10.1246/cl.160985. ISSN 0366-7022.
- ^ Switzer C, Moroney SE, Benner SA (1989). "Enzymatic incorporation of a new base of operations pair into Dna and RNA". J. Am. Chem. Soc. 111 (21): 8322–8323. doi:x.1021/ja00203a067.
- ^ a b Callaway E (May seven, 2014). "Scientists Create First Living Organism With 'Bogus' DNA". Nature News. Huffington Post. Retrieved viii May 2014.
- ^ Hirao I, Ohtsuki T, Fujiwara T, Mitsui T, Yokogawa T, Okuni T, Nakayama H, Takio Thousand, Yabuki T, Kigawa T, Kodama G, Yokogawa T, Nishikawa K, Yokoyama South (February 2002). "An unnatural base pair for incorporating amino acrid analogs into proteins". Nature Biotechnology. 20 (2): 177–82. doi:10.1038/nbt0202-177. PMID 11821864. S2CID 22055476.
- ^ Hirao I, Kimoto M, Mitsui T, Fujiwara T, Kawai R, Sato A, Harada Y, Yokoyama S (September 2006). "An unnatural hydrophobic base pair organization: site-specific incorporation of nucleotide analogs into Dna and RNA". Nature Methods. three (9): 729–35. doi:10.1038/nmeth915. PMID 16929319. S2CID 6494156.
- ^ Kimoto, Grand. et al. (2009) An unnatural base pair system for efficient PCR amplification and functionalization of DNA molecules. Nucleic acids Res. 37, e14
- ^ Yamashige R, Kimoto M, Takezawa Y, Sato A, Mitsui T, Yokoyama S, Hirao I (March 2012). "Highly specific unnatural base of operations pair systems equally a third base pair for PCR amplification". Nucleic Acids Inquiry. forty (6): 2793–806. doi:10.1093/nar/gkr1068. PMC3315302. PMID 22121213.
- ^ Kimoto K, Yamashige R, Matsunaga K, Yokoyama S, Hirao I (May 2013). "Generation of high-affinity DNA aptamers using an expanded genetic alphabet". Nature Biotechnology. 31 (5): 453–7. doi:10.1038/nbt.2556. PMID 23563318. S2CID 23329867.
- ^ a b c d Malyshev DA, Dhami K, Lavergne T, Chen T, Dai N, Foster JM, Corrêa IR, Romesberg FE (May 2014). "A semi-synthetic organism with an expanded genetic alphabet". Nature. 509 (7500): 385–8. Bibcode:2014Natur.509..385M. doi:ten.1038/nature13314. PMC4058825. PMID 24805238.
- ^ Sample I (May vii, 2014). "First life forms to pass on artificial DNA engineered by US scientists". The Guardian . Retrieved eight May 2014.
- ^ a b "Scientists create beginning living organism containing artificial Dna". The Wall Street Journal. Trick News. May 8, 2014. Retrieved eight May 2014.
- ^ a b Pollack A (May 7, 2014). "Scientists Add together Letters to Dna's Alphabet, Raising Promise and Fear". New York Times . Retrieved viii May 2014.
- ^ a b Evgenia N. Nikolova; Eunae Kim; Abigail A. Wise; Patrick J. O'Brien; Ioan Andricioaei; Hashim M. Al-Hashimi (2011). "Transient Hoogsteen base pairs in canonical duplex Deoxyribonucleic acid". Nature. 470 (7335): 498–502. Bibcode:2011Natur.470..498N. doi:10.1038/nature09775. PMC3074620. PMID 21270796.
- ^ Murphy, Frank V; Ramakrishnan, V (2004-11-21). "Construction of a purine-purine wobble base pair in the decoding center of the ribosome". Nature Structural & Molecular Biology. 11 (12): 1251–1252. doi:x.1038/nsmb866. ISSN 1545-9993. PMID 15558050. S2CID 27022506.
- ^ Vargas-Rodriguez, Oscar; Musier-Forsyth, Karin (June 2014). "Wobble puts RNA on target". Nature. 510 (7506): 480–481. doi:x.1038/nature13502. ISSN 1476-4687. PMID 24919145. S2CID 205239383.
- ^ Garg, Ankur; Heinemann, Udo (2017-xi-09). "A novel form of RNA double helix based on G·U and C·A+wobble base pairing". RNA. 24 (2): 209–218. doi:ten.1261/rna.064048.117. ISSN 1355-8382. PMC5769748. PMID 29122970.
- ^ Jun Aishima, Rossitza K. Gitti, Joyce Due east. Noah, Hin Hark Gan, Tamar Schlick, Cynthia Wolberger (2002). "A Hoogsteen base pair embedded in undistorted B‐DNA". Nucleic Acids Res. 30 (23): 5244–5252. doi:10.1093/nar/gkf661. PMC137974. PMID 12466549.
{{cite periodical}}: CS1 maint: multiple names: authors listing (link) - ^ a b Leontis, Neocles B; Westhof, Eric (2003). "Assay of RNA motifs". Current Stance in Structural Biology. 13 (three): 300–308. doi:10.1016/S0959-440X(03)00076-9. PMID 12831880.
- ^ Alberts B, Johnson A, Lewis J, Morgan D, Raff M, Roberts 1000, Walter P (December 2014). Molecular Biological science of the Prison cell (6th ed.). New York/Abingdon: Garland Scientific discipline, Taylor & Francis Group. p. 177. ISBN978-0-8153-4432-2.
- ^ "NIH ORDR – Glossary – C". Rarediseases.info.nih.gov. Retrieved 2012-07-16 .
- ^ Scott MP, Matsudaira P, Lodish H, Darnell J, Zipursky Fifty, Kaiser CA, Berk A, Krieger M (2004). Molecular Cell Biology (Fifth ed.). San Francisco: Westward. H. Freeman. p. 396. ISBN978-0-7167-4366-8.
...in humans 1 centimorgan on boilerplate represents a altitude of about seven.5x105 base of operations pairs.
Farther reading [edit]
- Watson JD, Baker TA, Bell SP, Gann A, Levine Yard, Losick R (2004). Molecular Biology of the Cistron (5th ed.). Pearson Benjamin Cummings: CSHL Press. (Run into esp. ch. 6 and nine)
- Sigel A, Sigel H, Sigel RK, eds. (2012). Interplay between Metal Ions and Nucleic Acids. Metal Ions in Life Sciences. Vol. 10. Springer. doi:10.1007/978-94-007-2172-2. ISBN978-nine-4007-2171-5. S2CID 92951134.
- Clever GH, Shionoya G (2012). "Chapter 10. Alternative DNA Base-Pairing through Metal Coordination". Interplay between Metal Ions and Nucleic Acids. Metal Ions in Life Sciences. Vol. x. pp. 269–294. doi:x.1007/978-94-007-2172-2_10. ISBN978-94-007-2171-v. PMID 22210343.
- Megger DA, Megger N, Mueller J (2012). "Affiliate 11. Metal-Mediated Base Pairs in Nucleic Acids with Purine and Pyrimidine-Derived Neucleosides". Coaction between Metal Ions and Nucleic Acids. Metal Ions in Life Sciences. Vol. ten. pp. 295–317. doi:10.1007/978-94-007-2172-2_11. ISBN978-94-007-2171-5. PMID 22210344.
External links [edit]
- DAN—webserver version of the EMBOSS tool for calculating melting temperatures

What Is Base Pair Rule,
Source: https://en.wikipedia.org/wiki/Base_pair
Posted by: glessnersopland.blogspot.com
0 Response to "What Is Base Pair Rule"
Post a Comment